
Page 6 - sarà_2002
P. 6
DDI_119.fm Page 6 Thursday, December 27, 2001 10:00 AM
6 M. Sarà and S. Morand
A 1998), as has its consequences in term of energy
used (Damuth, 1981).
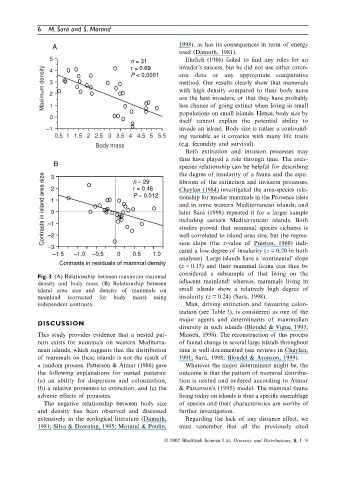
5 n = 31 invader’s success, but he did not use either exten-
Ehrlich (1986) failed to find any rules for an
Maximum density 3 P < 0.0001 sive data or any appropriate comparative
r = 0.69
4
method. Our results clearly show that mammals
with high density compared to their body mass
2
are the best invaders, or that they have probably
less chance of going extinct when living in small
1
populations on small islands. Hence, body size by
0
itself cannot explain the potential ability to
–1 invade an island. Body size is rather a confound-
0.5 1 1.5 2 2.5 3 3.5 4 4.5 5 5.5 ing variable as it covaries with many life traits
Body mass (e.g. fecundity and survival).
Both extinction and invasion processes may
thus have played a role through time. The area–
B n = 29 species relationship can be helpful for describing
Contrasts in island area size –1 2 1 0 r = 0.46 Cheylan (1984) investigated the area-species rela-
the degree of insularity of a fauna and the equi-
3
librium of the extinction and invasion processes.
P = 0.012
tionship for insular mammals in the Provence islets
and in some western Mediterranean islands; and
later Sarà (1998) repeated it for a larger sample
including eastern Mediterranean islands. Both
studies proved that mammal species richness is
–2
sion slope (the z-value of Preston, 1960) indi-
–3 well correlated to island area size, but the regres-
cated a low degree of insularity (z = 0.20 in both
–1.5 –1.0 –0.5 0 0.5 1.0
analyses). Large islands have a ‘continental’ slope
Contrasts in residuals of mammal density
(z = 0.13) and their mammal fauna can thus be
considered a subsample of that living on the
Fig. 3 (A) Relationship between maximum mammal
density and body mass. (B) Relationship between adjacent mainland; whereas, mammals living in
island area size and density of mammals on small islands show a relatively high degree of
mainland (corrected for body mass) using insularity (z = 0.24) (Sarà, 1998).
independent contrasts. Man, driving extinction and favouring colon-
ization (see Table 3), is considered as one of the
major agents and determinants of mammalian
DISCUSSION
diversity in such islands (Blondel & Vigne, 1993;
This study provides evidence that a nested pat- Masseti, 1998). The reconstruction of this process
tern exists for mammals on western Mediterra- of faunal change in several large islands throughout
nean islands, which suggests that the distribution time is well documented (see reviews in Cheylan,
of mammals on these islands is not the result of 1991; Sarà, 1998; Blondel & Aronson, 1999).
a random process. Patterson & Atmar (1986) gave Whatever the major determinant might be, the
the following explanations for nested patterns: outcome is that the pattern of mammal distribu-
(a) an ability for dispersion and colonization, tion is nested and ordered according to Atmar
(b) a relative proneness to extinction, and (c) the & Patterson’s (1995) model. The mammal fauna
adverse effects of parasites. living today on islands is thus a specific assemblage
The negative relationship between body size of species and their characteristics are worthy of
and density has been observed and discussed further investigation.
extensively in the ecological literature (Damuth, Regarding the lack of any distance effect, we
1981; Silva & Downing, 1995; Morand & Poulin, must remember that all the previously cited
© 2002 Blackwell Science Ltd, Diversity and Distributions, 8, 1–9