
Page 5 - Bracciali_2016
P. 5
120 C. Bracciali et al. / Marine Environmental Research 113 (2016) 116e123
0.0480
0.0465
0.0450
0.0435
0.0420
BCI 0.0405
0.0390
0.0375
LOW-HYDRO
0.0360 HIGH-HYDRO
< 50.00 50.00- 55.00- 60.00- 65.00- 70.00- > 75.00
54.95 59.95 64.95 69.95 74.95
SL class (mm)
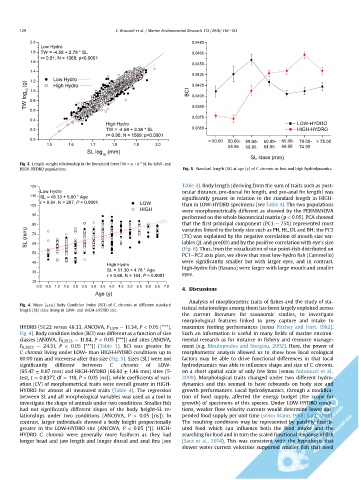
Fig. 3. Length-weight relationship in the linearized form TW ¼ a þ b * SL for LOW- and
HIGH-HYDRO populations. Fig. 5. Standard length (SL) at age (y) of C. chromis in low and high hydrodynamics.
120 Table 4). Body length (deriving from the sum of traits such as post-
Low Hydro ocular distance, pre-dorsal fin length, and pre-anal fin length) was
110 SL = 48.33 + 5.00 * Age significantly greater in relation to the standard length in HIGH-
r = 0.64, N = 207, P < 0.0001 LOW
100 than in LOW-HYDRO specimens (see Table 4). The two populations
HIGH were morphometrically different as showed by the PERMANOVA
90
performed on the whole biometrical matrix (p < 0.05). PCA showed
80 that the first principal component (PC1 ¼ 75%) represented most
SL (mm) 70 (7%) was explained by the negative correlation of mouth size var-
variables linked to the body size such as PH, HL, DL and BH; the PC2
iables (JL and preOD) and by the positive correlation with eye's size
60
(Fig. 6). Thus, from the visualization of our point-fish distributed on
50
PC1ePC2 axis plan, we show that most low-hydro fish (Cammello)
40 were significantly smaller but with larger eyes, and in contrast,
High Hydro
SL = 51.30 + 4.76 * Age high-hydro fish (Basana) were larger with large mouth and smaller
30
r = 0.68, N = 164, P < 0.0001 eyes.
20
0.0 0.5 1.0 1.5 2.0 2.5 3.0 3.5 4.0 4.5 5.0 5.5 6.0 6.5 7.0
4. Discussions
Age (y)
Analysis of morphometric traits of fishes and the study of sta-
Fig. 4. Mean (±s.e) Body Condition Index (BCI) of C. chromis at different standard
length (SL) class living in LOW- and HIGH-HYDRO site. tistical relationships among them has been largely exploited across
the current literature for taxonomic studies, to investigate
morphological features linked to prey capture and intake to
HYDRO (51.22 versus 48.33, ANCOVA, F 1,370 ¼ 11.34, P < 0.05 [***], maximize feeding performances (sensu Pitcher and Hart, 1982).
Fig. 4). Body condition index (BCI) was different as a function of size Such an information is useful in many fields of marine environ-
classes (ANOVA, F 6,2923 ¼ 11.84, P < 0.05 [***]) and sites (ANOVA, mental research as for instance in fishery and resource manage-
F 6,2923 ¼ 24.31, P < 0.05 [***]) (Table 3). BCI was greater for ment (e.g. Moutopoulos and Stergiou, 2002). Here, the power of
C. chromis living under LOW- than HIGH-HYDRO conditions up to morphometric analysis allowed us to show how local ecological
60.00 mm and viceversa after this size (Fig. 5). Sizes (SL) were not factors may be able to drive functional differences in that local
significantly different between C. chromis of LOW- hydrodynamics was able to influence shape and size of C. chromis,
(65.47 ± 0.97 mm) and HIGH-HYDRO (66.92 ± 1.46 mm) sites (T- on a short spatial scale of only few kms (sensu Antonucci et al.,
test, t ¼ 0.8377, df ¼ 118, P > 0.05 [ns]), while coefficients of vari- 2009). Morphological traits changed under two different hydro-
ation (CV) of morphometrical traits were overall greater in HIGH- dynamics and this seemed to have rebounds on body size and
HYDRO for almost all measured traits (Table 4). The regression growth performances. Local hydrodynamics, through a modifica-
between SL and all morphological variables was used as a tool to tion of food supply, affected the energy budget (the scope for
investigate the shape of animals under two conditions. Smaller fish growth) of specimens of this species. Under LOW-HYDRO condi-
had not significantly different slopes of the body height-SL re- tions, weaker flow velocity currents would determine lower sus-
lationships under two conditions (ANCOVA, P > 0.05 [ns]). In pended food supply per unit time (sensu Mann, 1988; Sar a, 2006).
contrast, larger individuals showed a body height proportionally The resulting conditions may be represented by patchily distrib-
greater in the LOW-HYDRO site (ANCOVA, P < 0.05 [*]). HIGH- uted food which can influence both the food intake and the
HYDRO C. chromis were generally more fusiform as they had searching for food and in turn the scaled functional response of fish
longer head and jaw length and longer dorsal and anal fins (see (Sar a et al., 2014). This was consistent with the hypothesis that
slower water current velocities supported smaller fish that need