
Page 4 - vol64-2011-215-222deidun
P. 4
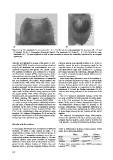
a) b)
Figure 2. a) The landmarks for pronotum (N = 4: 1-3-9-10) and the semi-landmarks for pronotum (N = 6) and
b) adopted for the P. bimaculata individuals studied. The landmarks for elytra (N = 4: 1-2-3-16) and the semi-
landmarks (N = 12) were digitized on half of each structure to remove the variability introduced by an eventual
asymmetry.
collected and digitized by means of the tpsDIG 2 soft- Phaleria species was reported (Deidun et al., 2010). A
ware (Rohlf, 2004). In order to better perform statistical possible reason for such a discrepancy might be the
analysis, the landmarks and semi-landmarks were suc- snapshot nature of the sampling conducted in the pre-
cessively recognized with tpsUtil 1.45 (Rohlf, 2008). sent study which might have resulted in P. acuminata
The resulting coordinates were subjected to a General- not being represented in collections, although there are
ized Procrustes Analysis (GPA), which removes all the no records of significant phenological differences be-
information unrelated to shape (Rohlf and Slice, 1990). tween the two species.
For the comparison of the configurations of landmarks Major body shape differences seem to be probably re-
and semi-landmarks, the Relative Warps Method was lated to geometric differences in elytra, as evident from
used (Bookstein, 1991; Rohlf, 1993). The relative warps deformation grids (figure 3): the elytra of the individu-
are principal component vectors of the partial warps, als from Vulcano (points 31-45 in the plot) were more
variables generated for thin-plate spline transformations elongated and elliptical in comparison to the Maltese
(Bookstein, 1989), and these were used to describe the individuals (1-15) and the Sicilian individuals (16-30),
major trends in shape variation among specimens within with the latter being positioned in the central area of the
the sample (Rohlf, 1993; 1996). Thin-plate spline de- plot.
formation grids were generated to facilitate description
of shape variation. The analyses were performed by The CVA showed a cumulative variance in the first
means of Relative Warps 1.39 software (Rohlf, 2004). two axis of 100%, for both pronotum and elytra analy-
sis. In the CVA scatterplot, the three groups, Vulcano,
In order to single out a putative relationship between Malta, Sicily, can be clearly distinguished (figure 4).
site and shape, a clustering of the studied specimens was The Mahalanobis distances (table 2) showed, in all
performed on the basis of their geographical position, pairwise comparisons, strong significant differences (p
through the Canonical Variate Analysis (CVA). The < 0.001) in body shape, highlighting the differentiation
CVA of the morphometric data was performed using of elytra-shape between the site Vulcano and the rest of
MorphoJ 1.01 software (Klingenberg, 2011) and used to samples.
simplify the identification of differences between prede-
fined group means (Zelditch et al., 2004). The observed inter-population shape differentiation
clustered the populations into three groups, according to
Results and discussion a spatial structure pattern, which could be described as
“site-specific”. A model of isolation by distance cannot
P. bimaculata individuals were recorded at 20 different
locations, of which 2 were located in Sicily, 9 in Table 2. Mahalanobis distances among groups. Below
circum-Sicilian islands and 9 in the Maltese Islands. In elytra, above pronotum analysis.
addition, three P. acuminata populations were also re-
corded along southern Sicilian shores (table 1). The two Malta Malta Sicily Vulcano
Phaleria species were never recorded in sympatry Sicily 3.1369*** 3.0108***
within the current study. Such an observation was unex- Vulcano 3.9024*** 3.1567***
pected for the Maltese beaches of Golden Bay and 6.8089*** 5.8927***
Mgiebah where a previous sympatry between the two
P-values from permutation tests (10000 permutation
rounds); ***P < 0.001.
218