
Page 42 - Lo Cascio_2015
P. 42
570 PIETRO LO CASCIO
Australasian islands (Fig. 7) constitute a clearly
distinct group from Lesser Sundas and Sulawesi
and are characterized by two main clusters: in the
first are included Maluku and the coastal islands
of western New Guinea (Biak, Misool, Roon,
Salawati, Umboi, Waigeo, Yapen), while in the
other are grouped New Guinea, its eastern satellites
(Baronga, Daru, Kiriwina, Lihir, Normanby,
Woodlark, Yule) and Bismarck Archipelago (New
Britain, New Ireland, New Hanover). The greater
faunistic affinity found between Papuan and
Bismarck islands is due to their geographical prox-
imity, but also to the fact that their faunas represent
fractions of the high diversity of New Guinea.
Within the Sunda Islands (Fig. 8), where some
islands (Sulawesi, Borneo, Java) host a large num-
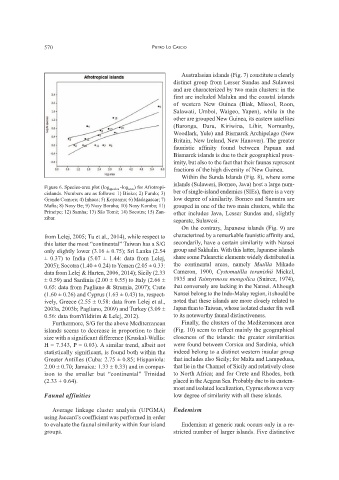
Figure 6. Species-area plot (log species -log area ) for Afrotropi-
cislands. Numbers are as follows: 1) Bioko; 2) Fundo; 3) ber of single-island endemics (SIEs), there is a very
Grande Comore; 4) Inhaca; 5) Koyaama; 6) Madagascar; 7) low degree of similarity. Borneo and Sumatra are
Mafia; 8) Nosy Be; 9) Nosy Boraha; 10) Nosy Komba; 11) grouped in one of the two main clusters, while the
Príncipe; 12) Samha; 13) São Tomé; 14) Socotra; 15) Zan- other includes Java, Lesser Sundas and, slightly
zibar.
separate, Sulawesi.
On the contrary, Japanese islands (Fig. 9) are
from Lelej, 2005; Tu et al., 2014), while respect to characterized by a remarkable faunistic affinity and,
this latter the most “continental” Taiwan has a S/G secondarily, have a certain similarity with Nansei
only slightly lower (3.16 ± 0.75); Sri Lanka (2.54 group and Sakhalin. With this latter, Japanese islands
± 0.37) to India (5.07 ± 1.44: data from Lelej, share some Palearctic elements widely distributed in
2005); Socotra (1.40 ± 0.24) to Yemen (2.05 ± 0.33: the continental areas, namely Mutilla Mikado
data from Lelej & Harten, 2006, 2014); Sicily (2.33 Cameron, 1900, Cystomutilla teranishii Mickel,
± 0.59) and Sardinia (2.00 ± 0.55) to Italy (2.66 ± 1935 and Taimyrmosa mongolica (Suárez, 1974),
0.65: data from Pagliano & Strumia, 2007); Crete that conversely are lacking in the Nansei. Although
(1.60 ± 0.26) and Cyprus (1.63 ± 0.43) to, respect- Nansei belong to the Indo-Malay region, it should be
ively, Greece (2.55 ± 0.58: data from Lelej et al., noted that these islands are more closely related to
2003a, 2003b; Pagliano, 2009) and Turkey (3.09 ± Japan than to Taiwan, whose isolated cluster fits well
0.56: data fromYildirim & Lelej, 2012). to its noteworthy faunal distinctiveness.
Furthermore, S/G for the above Mediterranean Finally, the clusters of the Mediterranean area
islands seems to decrease in proportion to their (Fig. 10) seem to reflect mainly the geographical
size with a significant difference (Kruskal-Wallis: closeness of the islands: the greater similarities
H = 7.343, P = 0.03). A similar trend, albeit not were found between Corsica and Sardinia, which
statistically significant, is found both within the indeed belong to a distinct western insular group
Greater Antilles (Cuba: 2.75 ± 0.85; Hispaniola: that includes also Sicily; for Malta and Lampedusa,
2.00 ± 0.70; Jamaica: 1.33 ± 0.33) and in compar- that lie in the Channel of Sicily and relatively close
ison to the smaller but “continental” Trinidad to North Africa; and for Crete and Rhodes, both
(2.33 ± 0.64). placed in the Aegean Sea. Probably due to its eastern-
most and isolated localization, Cyprus shows a very
Faunal affinities low degree of similarity with all these islands.
Average linkage cluster analysis (UPGMA) Endemism
using Jaccard’s coefficient was performed in order
to evaluate the faunal similarity within four island Endemism at generic rank occurs only in a re-
groups. stricted number of larger islands. Five distinctive