
Page 5 - THE_ISLAND_RULE_2012
P. 5
THE ISLAND RULE IN LARGE MAMMALS 1735
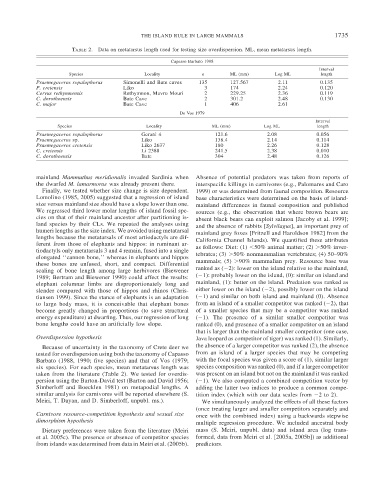
TABLE 2. Data on metatarsus length used for testing size overdispersion. ML, mean metatarsus length.
Capasso Barbato 1988
Interval
Species Locality n ML (mm) Log ML length
Praemegaceros ropalophorus Simonelli and Bate caves 135 127.567 2.11 0.135
P. cretensis Liko 3 174 2.24 0.120
Cervus rethymnensis Rethymnon, Mavro Mouri 2 229.25 2.36 0.119
C. dorothoensis Bate Cave 2 301.2 2.48 0.130
C. major Bate Cave 1 406 2.61
De Vos 1979
Interval
Species Locality ML (mm) Log ML length
Praemegaceros ropalophorus Gerani 4 121.6 2.08 0.056
Praemegaceros sp. Liko 138.4 2.14 0.114
Praemegaceros cretensis Liko 2637 180 2.26 0.128
C. cretensis Li 2388 241.5 2.38 0.010
C. dorothoensis Bate 304 2.48 0.126
mainland Mammuthus meridionalis invaded Sardinia when Absence of potential predators was taken from reports of
the dwarfed M. lamarmorae was already present there. interspecific killings in carnivores (e.g., Palomares and Caro
Finally, we tested whether size change is size dependent. 1999) or was determined from faunal composition. Resource
Lomolino (1985, 2005) suggested that a regression of island base characteristics were determined on the basis of island-
size versus mainland size should have a slope lower than one. mainland differences in faunal composition and published
We regressed third lower molar lengths of island fossil spe- sources (e.g., the observation that where brown bears are
cies on that of their mainland ancestor after partitioning is- absent black bears can exploit salmon [Jacoby et al. 1999];
land species by their CLs. We repeated the analyses using
and the absence of rabbits [Sylvilagus], an important prey of
humeri lengths as the size index. We avoided using metatarsal
mainland grey foxes [Fritzell and Haroldson 1982] from the
lengths because the metatarsals of most artiodactyls are dif-
California Channel Islands). We quantified these attributes
ferent from those of elephants and hippos: in ruminant ar-
as follows: Diet: (1) 50% animal matter; (2) 50% inver-
tiodactyls only metatarsals 3 and 4 remain, fused into a single
tebrates; (3) 50% nonmammalian vertebrates; (4) 50–90%
elongated ‘‘cannon bone,’’ whereas in elephants and hippos
mammals; (5) 90% mammalian prey. Resource base was
these bones are unfused, short, and compact. Differential
ranked as ( 2): lower on the island relative to the mainland,
scaling of bone length among large herbivores (Biewener
( 1): probably lower on the island, (0): similar on island and
1989; Bertram and Biewener 1990) could affect the results:
elephant columnar limbs are disproportionately long and mainland, (1): better on the island. Predation was ranked as
slender compared with those of hippos and rhinos (Chris- either lower on the island ( 2), possibly lower on the island
tiansen 1999). Since the stance of elephants is an adaptation ( 1) and similar on both island and mainland (0). Absence
to large body mass, it is conceivable that elephant bones from an island of a smaller competitor was ranked ( 2), that
become greatly changed in proportions (to save structural of a smaller species that may be a competitor was ranked
energy expenditure) at dwarfing. Thus, our regression of long ( 1). The presence of a similar smaller competitor was
bone lengths could have an artificially low slope. ranked (0), and presence of a smaller competitor on an island
that is larger than the mainland smaller competitor (one case,
Overdispersion hypothesis Java leopard as competitor of tiger) was ranked (1). Similarly,
Because of uncertainty in the taxonomy of Crete deer we the absence of a larger competitor was ranked (2), the absence
tested for overdispersion using both the taxonomy of Capasso from an island of a larger species that may be competing
Barbato (1988, 1990; five species) and that of Vos (1979; with the focal species was given a score of (1), similar larger
six species). For each species, mean metatarsus length was species composition was ranked (0), and if a larger competitor
taken from the literature (Table 2). We tested for overdis- was present on an island but not on the mainland it was ranked
persion using the Barton-David test (Barton and David 1956; ( 1). We also computed a combined competition vector by
Simberloff and Boecklen 1981) on metapodial lengths. A adding the latter two indices to produce a common compe-
similar analysis for carnivores will be reported elsewhere (S. tition index (which with our data scales from 2to2).
Meiri, T. Dayan, and D. Simberloff, unpubl. ms.). We simultaneously analyzed the effects of all these factors
(once treating larger and smaller competitors separately and
Carnivore resource-competition hypothesis and sexual size once with the combined index) using a backwards stepwise
dimorphism hypothesis
multiple regression procedure. We included ancestral body
Dietary preferences were taken from the literature (Meiri mass (S. Meiri, unpubl. data) and island area (log trans-
et al. 2005c). The presence or absence of competitor species formed, data from Meiri et al. [2005a, 2005b]) as additional
from islands was determined from data in Meiri et al. (2005b). predictors.